早上被闹钟叫醒的那一刻,大脑实际上经历了一次明显的状态切换,从睡眠到清醒,从“关”到“开”。
日常生活中,人始终处在这类感觉信息的包围之中。闹钟声、光线变化、有人拍你肩膀,这些外界刺激都能瞬间改变你的脑状态。但问题是,它们是怎么做到的?为什么同一个声音,有时候可以把人从深度睡眠中唤醒,有时候却被大脑自动屏蔽?为什么有些人听见门铃响就心跳加速,有些人却无感?
这些现象指向一个更底层的问题:感觉信息是如何调控“唤醒”(arousal)这一核心脑功能的?
围绕这一问题,清华大学生物医学工程学院苑克鑫团队受邀在神经科学旗舰综述期刊《Annual Review of Neuroscience》发表年度综述,为该问题的解答提出了理论框架。
点击此处查看更多论文信息
1 唤醒这件事,得拆开了看
传统研究通常将“唤醒”作为整体状态来处理,可以简单理解为从睡眠到清醒的状态变化。但这篇综述提出有必要对这一概念进行拆分,即分为一般性唤醒(general arousal)与特定唤醒(specific arousal)。
一般性唤醒是全局的,例如从睡眠到清醒的切换,决定大脑是“开机”还是“关机”。 特定唤醒则具有行为指向——看见蛇时的心跳加速、听见婴儿哭声时的警觉、闻到食物气味时的反应,都是在“开机”状态下,大脑根据环境需求调用的不同模式。
这一划分的关键在于,“开机”只是前提,更关键的是“用什么模式开机”。 不同情境下,大脑会调动不同的反应方式:例如在野外面对风吹草动时进入防御模式;而在家里闻到厨房香味则进入觅食模式。同一唤醒状态下可以调用不同的行为输出,这种灵活性正构成了生物智能的核心。
2 三个系统,三种逻辑
在机制层面,综述将感觉诱发唤醒的神经基础归纳为三类系统,它们的作用方式存在明显差异。
第一个是感觉系统本身,尤其是高阶感觉丘脑(图1)。传统观点通常将丘脑视为感觉信息的中转站——声音经由耳部传入丘脑,再传到皮层。但该综述的证据表明,高阶感觉丘脑并非被动中转站,而是能够对刺激的显著性(saliency)进行评估,并决定是否唤醒皮层,及用哪种方式唤醒。
以听觉为例,听觉丘脑内侧部(ATm)被证明是多模态感觉诱发唤醒的枢纽,不仅处理声音,还接收视觉、躯体感觉的输入,并且直接往下丘脑投射。这意味着,在感觉信息尚未传至皮层之前,丘脑就已经开始影响大脑的状态选择。
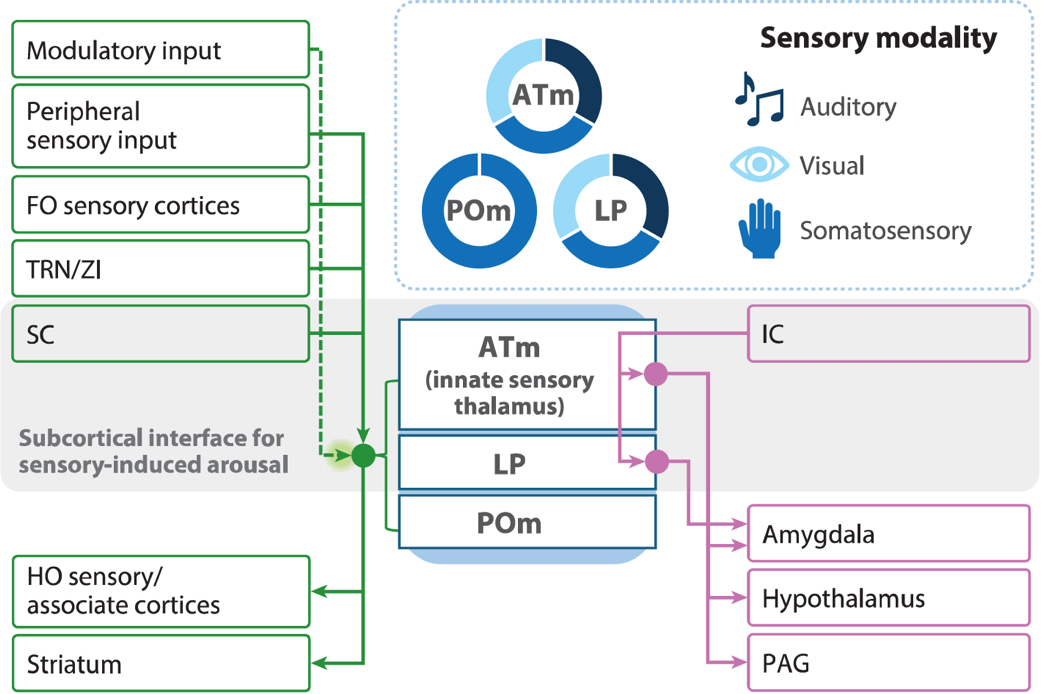
图1 高阶感觉丘脑介导多模态感觉诱发唤醒的神经基础
第二个是一般性唤醒系统,即脑干中的神经调质核团,如蓝斑、中缝背核与腹侧被盖区。传统上,这些结构对感觉信息的高度敏感,被认为能够调控睡眠-清醒。综述则指出,唤醒效果并不完全由刺激本身决定,例如蓝斑核去甲肾上腺素能神经元在感觉诱发唤醒中的作用,主要依赖其自身的“基线活动”水平,而非被声音诱发的瞬时放电。
这解释了一个日常现象:同样的声音,在深度睡眠时叫不醒你,而在睡眠浅的时候却可以,关键差异并不在于刺激,而在于大脑自身的状态。
第三个是与特定行为相关的功能区,包括下丘脑室旁核、腹内侧下丘脑及终纹床核等。这些区域对感觉信息具有极高的选择性。例如,腹内侧下丘脑的SF1神经元对捕食者相关刺激具有持续反应,而对纯音或食物的刺激则几乎不响应。同时,这种神经活动具有持续性,即使刺激消失后仍然保持活跃,让你在危险过去之后仍然保持警惕。
3 三个系统如何互相配合?
把这三大系统的反应曲线放在同一框架下,可以更清晰地理解其分工(图2)。
特定功能系统(橙色)对刺激显著性的的响应近乎“全或无”:对无关刺激几乎不响应,一旦超过阈值即被激活,诱发特定唤醒状态,如同是一个高分辨率的问题处理专家。相比之下,一般性唤醒系统(蓝色)的响应更为平缓,以连续方式调节整体唤醒水平。感觉系统(绿色)则与刺激显著性呈近似线性关系,其输出在前两者基础上叠加,参与调控最终的唤醒状态。
这三大系统在不同“分辨率”下对刺激进行评估,当它们的输出叠加,则能够共同决定个体最终进入的状态。该关系可表示为:
全局唤醒 = f₁(S) + f₂(S) + f₃(S)
其中,S表示感觉显著性,但并非单一维度,不同系统对其不同维度具有不同敏感性,并各自存在响应最大化的“偏好维度”。这一框架对应三种分工明确的调控模式。当在不同个体或不同状态下,各系统的基线水平与响应增益存在差异,即使相同刺激也可能产生不同效果。

图2 由感觉刺激诱发唤醒水平变化的机制示意图,展示了特定功能系统(橙色)、感觉系统(绿色)和一般性唤醒系统(蓝色)的调控作用。
4 唤醒与情感:一枚硬币的两面
上述框架下更进一步的问题是:情感与唤醒之间有何关联?直觉告诉我们,它们密切相关。害怕时心跳加速,开心时精力充沛,抑郁时什么都提不起劲。但这一相关性是否拥有更结构化里的联系,仍需进一步解释。
这篇综述提出,情感状态可以理解为特定唤醒模式的持续表达。具体而言,焦虑是防御相关唤醒系统的持续高增益状态;抑郁表现为一般性唤醒水平降低,同时正向行为(觅食、社交)相关的特定唤醒系统受到抑制;创伤后应激障碍则体现为特定唤醒状态在刺激消失后仍持续存在,难以及时关闭。
由此,情感障碍可能源于唤醒调控系统的功能失衡,并非“醒着”或“睡着”的宏观状态出了问题,而是“用什么模式醒着”这种精细调控出了问题。
从这个角度看,感觉信息之所以能影响情感,是因为感觉信息是唤醒系统的“遥控器”。不同参数的光、声、躯体感觉刺激,能选择性地激活或抑制不同的唤醒子系统,从而改变大脑的状态模式。这一机制也为基于感觉刺激的情感干预提供了神经基础。
5 仍待回答的关键问题
综述最后指出,该领域仍有若干关键问题有待进一步梳理:不同唤醒状态究竟由并行通路介导,还是汇聚到共同枢纽?特定功能系统的脑区内部是否存在专门的“唤醒神经元”? 能否建立刺激参数-神经活动-行为概率的量化关系?这些问题仍是推动相关研究继续深入的关键方向。
更为重要的是,大脑如何主动与外界环境进行交互的议题中,现有的预测编码框架通常将感觉结构当成传递预测误差的中转站。但综述梳理的证据表明,高阶感觉丘脑本身也在主动整合唤醒信号,且这一过程并不依赖皮层输入。这意味着,未来的计算模型或许需要重新评估这些传统上被视为执行基础功能的脑区,进一步考量其在唤醒调控中的功能复杂性。
原文链接:https://doi.org/10.1146/annurev-neuro-112723-034728



 清华大学医学科学楼(清华大学西北门南行50米路西侧)
清华大学医学科学楼(清华大学西北门南行50米路西侧) 62787861
62787861  sygcxybgs@mail.tsinghua.edu.cn
sygcxybgs@mail.tsinghua.edu.cn 微信公众号
微信公众号
